В 1945 г. Дж. Бидлом и Э. Татумом была сформулирована гипотеза, которую можно выразить формулой "Один ген - один фермент". Согласно этой гипотезе, каждая стадия метаболического процесса, приводящая к образованию в организме (клетке) какого-то продукта, катализируется белком-ферментом, за синтез которого отвечает один ген.
Позднее было показано, что многие белки имеют четвертичную структуру, в образовании которой принимают участие разные пептидные цепи. Поэтому формула, отражающая связь между геном и признаком, была несколько преобразована: "Один ген - один полипептид".
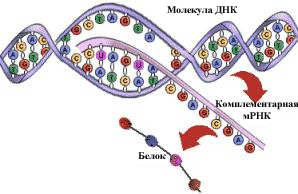
Изучение химической организации Э. Чаргаффом наследственного материала и процесса реализации генетической информации привело к формированию представления о гене как о фрагменте молекулы ДНК, транскрибирующемся в виде молекулы РНК, которая кодирует аминокислотную последовательность пептида или имеет самостоятельное значение (тРНК и рРНК).
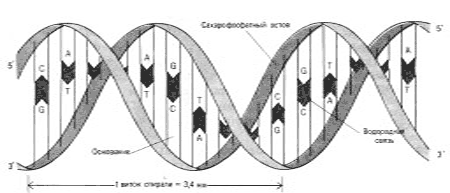
Также ценные сведения о структуре ДНК дали результаты рентгеноструктурного анализа. Рентгеновские лучи, проходя через кристалл ДНК, претерпевают дифракцию, т.е. отклоняются в определенных направлениях. Степень и характер отклонения зависят от структуры самой молекулы. Анализ дифракционных рентгенограмм привел к заключению, что азотистые основания уложены на подобие стопки тарелок. Рентгенограммы позволили выявить в ДНК 3 главных периода: 0,34, 2 и 3,4, которые оказались размерами в модели ДНК, предложенной Дж.Уотсоном и Ф.Криком. 0,34 нм - расстояние между последовательными нуклеотидами, 2 нм - толщина цепи, 3,4 нм - расстояние между последовательными витками спирали.
В конце двадцатых годов советские генетики А. С. Серебровский и Н. П. Дубинин экспериментально показали, что ген не является единицей мутации, что он имеет сложную структуру: состоит из нескольких субъединиц, способных самостоятельно мутировать (ступенчатый аллелизм, или центровая теория гена). Весь ген (базиген) может состоять из отдельных центров, трансгенов, каждый из которых несет сходную функцию. Мутация может нарушать деятельность одного из трансгенов, не затрагивая других.
Несколько позже идея о сложном строении гена была подкреплена экспериментами по внутригенному кроссинговеру на дрозофиле по локусам lozenge, white и др. (работы Э. Льюиса, М. Грина и др.).
Таким образом, к 1950 году ген представлялся как участок хромосомы, контролирующий развитие определенного признака, имеющий определенную линейную протяженность и способный мутировать в разных участках и быть разделенным кроссинговером. Ген комплексен, так как его отдельные участки могут различаться по функциям, и в их совместной деятельности существует определенная субординация.